シェア:



アンチコドンの説明、機能、およびコドンとの違い
A アンチコドン その機能は、メッセンジャーRNA分子(mRNA)中に存在する3ヌクレオチドの別の配列を認識することである、トランスファーRNA分子(tRNA)中に存在する3ヌクレオチドの配列である。.
コドンとアンチコドンの間のこの認識は逆平行です。すなわち、一方は5 ' - > 3'方向にあり、もう一方は3 ' - > 5'方向にあります。 3つのヌクレオチド(トリプレット)の配列間のこの認識は翻訳過程の基本である。すなわち、リボソーム内のタンパク質の合成において.
したがって、翻訳中に、メッセンジャーRNA分子は、トランスファーRNAのアンチコドンによるそれらのコドンの認識を通して「読み取られる」。これらの分子は、リボソームで形成されているタンパク質分子に特定のアミノ酸を移動させるため、このように呼ばれています。.
20個のアミノ酸があり、それぞれ特定のトリプレットでコードされています。しかしながら、いくつかのアミノ酸は複数のトリプレットによってコードされている.
さらに、いくつかのコドンは、アミノ酸が結合していない転移RNA分子中のアンチコドンによって認識される。これらはいわゆる終止コドンです。.
索引
- 1説明
- 2つの機能
- 3アンチコドンとコドンの違い
- 4ローリングの仮説
- 4.1 RNAとアミノ酸
- 5参考文献
説明
アンチコドンは、以下の含窒素塩基:アデニン(A)、グアニン(G)、ウラシル(U)またはシトシン(C)のいずれかを含むことができる3つのヌクレオチドの配列によって形成される。それはコードのように動作します.
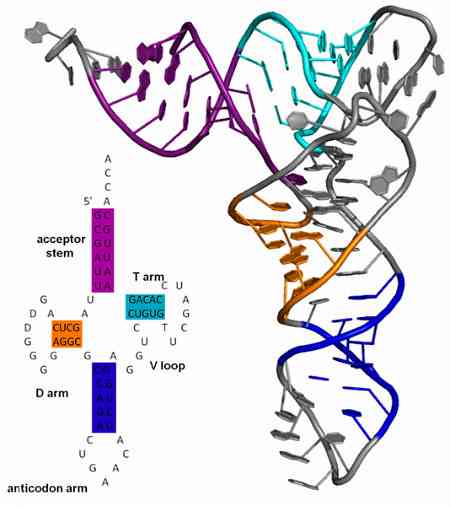
アンチコドンは常にトランスファーRNA分子に見られ、常に3 '→5'方向にある。これらのtRNAの構造はクローバーに似ていますが、4つのループ(または複数のループ)に分割されています。ループの1つにアンチコドンがあります.
アンチコドンは、メッセンジャーRNAのコドンの認識、そしてその結果としてすべての生細胞におけるタンパク質合成の過程に不可欠です。.
機能
アンチコドンの主な機能は、メッセンジャーRNA分子中のコドンを形成するトリプレットの特異的認識である。これらのコドンは、タンパク質中のアミノ酸の順序を決定するためにDNA分子からコピーされた指示です。.
転写(メッセンジャーRNAコピーの合成)は5 '→3'方向に起こるので、メッセンジャーRNA中のコドンはこの配向を有する。それ故、トランスファーRNA分子中に存在するアンチコドンは反対方向、3 '→5'を有さなければならない。.
この連合は相補性によるものです。例えば、1つのコドンが5'-AGG-3 'の場合、アンチコドンは3'-UCC-5'です。コドンとアンチコドンとの間のこの種の特異的相互作用は、メッセンジャーRNA中のヌクレオチド配列がタンパク質内のアミノ酸配列をコードすることを可能にする重要な工程である。.
アンチコドンとコドンの違い
- アンチコドンはtRNA中のトリヌクレオチド単位であり、mRNA中のコドンに相補的である。それらは、タンパク質産生中にtRNAが正しいアミノ酸を送達することを可能にする。対照的に、コドンは、タンパク質合成において特定のアミノ酸をコードするDNAまたはmRNAにおけるトリヌクレオチドの単位である。.
- アンチコドンは、mRNAのヌクレオチド配列とタンパク質のアミノ酸配列との間の結合である。逆に、コドンは、DNAがある核からタンパク質合成が起こるリボソームへ遺伝情報を伝達する。.
- アンチコドンは、DNAおよびmRNA分子に位置するコドンとは異なり、tRNA分子のアンチコドンアームにあります。.
- アンチコドンはそれぞれのコドンに相補的である。対照的に、mRNAのコドンはDNAの特定の遺伝子のヌクレオチドのトリプレットに相補的です.
- tRNAはアンチコドンを含む。対照的に、mRNAは多数のコドンを含む。.
ローリング仮説
均衡仮説は、メッセンジャーRNAのコドンの3番目のヌクレオチドとトランスファーRNAのアンチコドンの1番目のヌクレオチドとの間の接合部は、トリプレットの他の2つのヌクレオチド間の接合部よりも特異性が低いことを提案する。.
クリックは、この現象を各コドンの3番目の位置の「揺れ」として説明した。その位置で何かが起こり、それによって共用体は通常よりも厳格ではなくなります。それはまた、ウォブリングまたはタンボレオとしても知られています。.
このクリックウォブル仮説は、与えられたtRNAのアンチコドンが2つまたは3つの異なるmRNAコドンとどのように対になることができるかを説明しています.
Crickは、塩基対(tRNAのアンチコドンの塩基59とmRNAのコドンの塩基39との間)は通常より厳密ではないので、この部位では一定の「ゆらぎ」または低い親和性が許されると提案した。.
結果として、単一のtRNAはしばしば、与えられたアミノ酸を特定する2つまたは3つの関連コドンを認識します。.
通常、tRNAアンチコドンの塩基とmRNAコドンとの間の水素結合は、コドンの最初の2塩基についてのみ塩基対合の厳密な規則に従う。しかしながら、この効果は全てのmRNAコドンの全ての3番目の位置では起こらない。.
RNAとアミノ酸
ぐらつき仮説に基づいて、完全な変性を示すコドンを有する各アミノ酸について少なくとも2つのトランスファーRNAの存在が予測され、これは真実であることが証明されている。.
この仮説はまた、6つすべてのセリンコドンについて3つのトランスファーRNAの出現を予測した。実際、セリンに対する3つのtRNAが特徴付けられている。
- セリン1のTRNA(アンチコドンAGG)はコドンUCUおよびUCCに結合する.
- セリン2(アンチコドンAGU)のTRNAはコドンUCAおよびUCGに結合する.
- セリン3のTRNA(アンチコドンUCG)はコドンAGUおよびAGCに結合する.
これらの特異性は、インビトロでの精製アミノアシルtRNAトリヌクレオチドのリボソームへの刺激された結合によって検証された。.
最後に、いくつかのトランスファーRNAはヒポキサンチンプリンから作られるイノシン塩基を含んでいます。イノシンはアデノシンの転写後修飾によって産生される.
Crick wobble仮説は、イノシンがアンチコドンの5 '末端(振動位置)に存在する場合、それはコドンでウラシル、シトシンまたはアデニンと対になると予測した.
実際、アンチコドンの5 '位にイノシン(I)を含む精製アラニルtRNAは、GCU、GCCまたはGCAのトリヌクレオチドで活性化されたリボソームに結合する。.
アンチコドンの5 '位をイノシンで精製した他のtRNAでも同じ結果が得られた。したがって、クリックのゆらぎ仮説は、tRNAとコドンの間の関係を非常によく説明しています。.
参考文献
- Brooker、R.(2012). 遺伝学の概念 (第1版)。マッグロウヒル会社.
- Brown、T.(2006). ゲノム3 (3第二)ガーランドサイエンス.
- グリフィス、A。、ウェスラー、S、キャロル、S&Doebley、J(2015). 遺伝子解析の紹介 (第11版)。 W.H.フリーマン
- ルイス、R.(2015). ヒト遺伝学:概念と応用(第11版)。マッグロウヒル教育.
- Snustad、D.&Simmons、M.(2011). 遺伝学の原則(第6版)。ジョンワイリーと息子.