シェア:



レプリケーションフォークとは何ですか?
の レプリケーションフォーク それはDNA複製が起こる点であり、それは成長点とも呼ばれる。それはY字形をしており、そして複製が進むにつれて、ヘアピンはDNA分子によって置き換えられる。.
DNA複製は、細胞内の遺伝物質の複製を伴う細胞プロセスです。 DNAの構造は二重らせんであり、その内容を複製するためにはそれを開かなければならない。複製は半保存的プロセスであるため、各鎖は新しいDNA鎖の一部になります。.
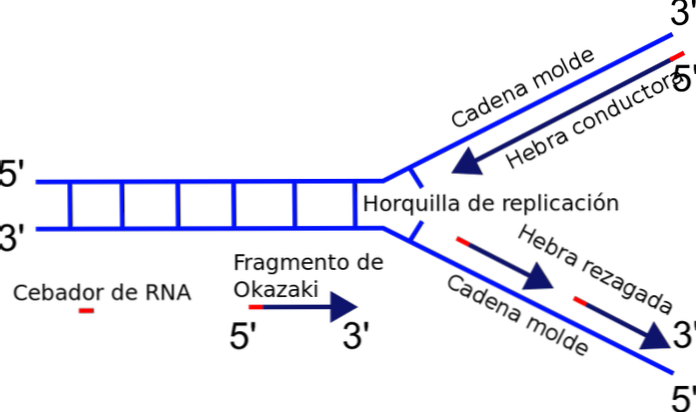
複製フォークは、新たに分離された鋳型または鋳型鎖とまだ複製されていない二本鎖DNAとの間の接合部の間にちょうど形成される。 DNA複製を開始すると、一方の鎖が容易に複製され、もう一方の鎖は極性の問題に直面します。.
鎖の重合を担う酵素 - DNAポリメラーゼ - は、5'-3 '方向にのみDNA鎖を合成します。したがって、一方の鎖は連続的であり、他方の鎖は不連続な複製を受け、岡崎の断片を生成する。.
索引
- 1 DNAの複製と複製フォーク
- 1.1単方向レプリケーションと双方向レプリケーション
- 関与する酵素
- 1.3複製開始とフォーク形成
- 1.4フォークの伸びと動き
- 1.5終了
- 2 DNA複製は半保存的
- 3極性の問題
- 3.1ポリメラーゼのしくみ?
- 3.2岡崎破片の製造
- 4参考文献
DNAの複製と複製フォーク
DNAは、いくつかのウイルスを除いて、すべての生物の必要な遺伝情報を保持する分子です。.
4つの異なるヌクレオチド(A、T、G、C)からなるこの巨大なポリマーは、真核生物の核内に存在し、これらの人間の組織を構成する各細胞には存在しません。コアの).
細胞が分裂するたびに、遺伝物質を持つ娘細胞を生み出すためにDNAを複製しなければなりません.
単方向および双方向のレプリケーション
複製は、始点での複製フォークの形成に応じて、単方向または双方向になります。.
論理的には、一方向の複製の場合は1つのフォークのみが形成され、双方向複製の場合は2つのフォークが形成されます。.
関与する酵素
この過程のためには、複雑な酵素機構が必要であり、それは素早く作用し、そして正確にDNAを複製することができる。最も重要な酵素は、DNAポリメラーゼ、DNAプライマーゼ、DNAヘリカーゼ、DNAリガーゼおよびトポイソメラーゼです。.
複製の開始とフォークの形成
DNA複製は分子内のランダムな場所からは開始されません。複製開始点を示すDNA内の特定の領域があります.
ほとんどのバクテリアでは、バクテリアの染色体は単一のATに富む出発点を持っています。この構成は論理的です、なぜならそれは領域の開放を容易にするからです(ATペアは2つの水素橋によって結合されますが、GCペアは3つによって結合されます)。.
DNAが開き始めると、Y字型の構造が形成されます。複製フォーク.
フォークの伸びと動き
DNAポリメラーゼは娘鎖の合成を一から始めることはできない。ポリメラーゼがどこから重合し始めるかを持つために3 '末端を持つ分子が必要です。.
この遊離3 '末端は、プライマーまたはプライマーと呼ばれる小分子ヌクレオチドによって提供される。最初のものはポリメラーゼの一種のフックとして機能します.
複製の過程で、複製フォークはDNAに沿って移動する能力を持ちます。複製フォークのピッチは二重バンド娘分子の形成を指示する二つの単一バンドDNA分子を残す.
DNA分子を広げるヘリカーゼ酵素の働きでフォークは前進することができます。この酵素は塩基対間の水素結合を切断し、フォークの移動を可能にします.
終了
2つのフォークが起点から180°Cになると、複製は終了します。.
この場合、我々はバクテリアの複製過程がどのように流れるかについて話します、そしてそれは複製を含む環状分子の全体のねじれ過程を強調することが必要です。トポイソメラーゼは分子の巻き戻しに重要な役割を果たす.
DNA複製は半保存的です
あなたはDNA中でどのように複製が起こるのか疑問に思ったことがありますか?つまり、二重らせんから別の二重らせんが生じるはずですが、どうすればよいのでしょうか。数年間、これは生物学者の間では未解決の問題でした。 2本の古いストランドと2本の新しいストランド、または2本の螺旋を形成するための新しいスレッドと古いストランドの組み合わせ.
1957年に、この質問は研究者Matthew MeselsonとFranklin Stahlによって答えられました。著者によって提案された複製モデルは半保存的であった.
MeselsonとStahlは、複製の結果は2本の二本鎖DNA分子であると述べた。得られた分子のそれぞれは、古い鎖(母親または最初の分子からのもの)および新しく合成された新しい鎖からなる。.
極性の問題
ポリメラーゼのしくみ?
DNAらせんは、逆平行に走る2本の鎖で形成されます。1本は5'-3 '方向に、もう1本は3'-5'.
複製過程において最も重要な酵素はDNAポリメラーゼであり、これは鎖に付加されるであろう新しいヌクレオチドの結合を触媒することに関与している。 DNAポリメラーゼは鎖を5'− 3 '方向にのみ伸長することができる。この事実は、複製フォーク内のチェーンの同時複製を妨げる.
なんで?ヌクレオチドの付加は、ヒドロキシル基(−OH)が見出される3 'の遊離末端で起こる。したがって、3 '末端へのヌクレオチドの末端付加によって、鎖のうちの一方だけが容易に増幅され得る。これは導電性または連続ストランドと呼ばれます.
岡崎破片の製造
他方の鎖は伸長することができない。なぜなら、遊離末端は5 'であり、3'ではなく、そしてポリメラーゼは5 '末端へのヌクレオチドの付加を触媒しないからである。問題は、それぞれが5 'から3'への複製の通常の方向にある複数の短い断片(130から200ヌクレオチド)の合成によって解決される。.
断片のこの不連続合成は、各部分の結合、DNAリガーゼによって触媒される反応で終わる。このメカニズムの発見者である岡崎玲二を記念して、合成された小さなセグメントは岡崎の破片と呼ばれます.
参考文献
- Alberts、B.、Bray、D.、Hopkin、K.、Johnson、A.D.、Lewis、J.、Raff、M.、...&Walter、P.(2015). 必須細胞生物学. ガーランドサイエンス.
- Cann、I. K.&Ishino、Y.(1999)。古細菌DNA複製:パズルを解くための断片の同定. 遺伝学, 152(4)、1249〜67.
- クーパー、G。M。&ハウスマン、R。E。(2004). 細胞:分子アプローチ. Medicinska naklada.
- Garcia-Diaz、M.、&Bebenek、K.(2007)。 DNAポリメラーゼの多機能. 植物科学における批評的レビュー, 26(2)、105〜122.
- Lewin、B。(2008). 遺伝子IX. Mc Graw-Hill Interamericana.
- Shcherbakova、P。V.、Bebenek、K。、およびKunkel、T。A.(2003)。真核生物DNAポリメラーゼの機能. サイエンスのSAGE KE, 2003年(8)、3.
- Steitz、T. A.(1999)。 DNAポリメラーゼ:構造的多様性と共通のメカニズム. バイオケミストリージャーナル, 274(25)、17395〜17398.
- Watson、J. D.(2006). 遺伝子の分子生物学. 編集Panamericana Medical.
- Wu、S.、Beard、W. A.、Pedersen、L. G.、&Wilson、S. H.(2013)。 DNAポリメラーゼ構造の構造比較はポリメラーゼ活性部位へのヌクレオチドゲートウェイを示唆する. 化学レビュー, 114(5)、2759-74.