シェア:



有孔虫の特徴、形態、分類、ライフサイクル
の 有孔虫 それらはアメーバ原生動物、いくつかの海洋および他の淡水のグループです。彼らは第一次時代(カンブリア紀)の初めに現れ、彼らの後継者たちは現在の海洋に居住しています。それらは沿岸地域(低または高塩分)から海底まで、そして熱帯地方から寒い北極海と南極海まで見つけることができます.
その分布はいくつかの要因に依存します。例えば、大きく頻繁な気温変化を支える種がありますが、他のものはそれを乗り切ることができませんでした。そのため、海洋の熱構造は有孔虫群の間の重要な違いを示しています。.

同様に、深さもまた、それらが光の透過に直接入射するために、有孔虫の分布における決定的要因である。一方、圧力は以前の要因(温度と深さ)に関係しており、COの溶解度に直接影響します。2, 貝殻の形成のための炭酸カルシウムの分泌に影響するもの.
一方、浅い地域の水のエネルギーは、それが基質の種類(硬いまたは柔らかい)と栄養素の分布に影響を与えるので、関連する要素です。.
同様に、塩分濃度、水の濁度、pH、微量元素や有機成分の存在、流れ、沈降速度などの他の要因が、有孔虫の分布を局所的に決定する可能性があります。.
索引
- 1知られている種
- 2つの特徴
- 2.1有孔虫の大きさと形態に関わる要因
- 3分類
- 4分類
- 4.1アタラメア
- 4.2モノタラメア
- 4.3 Xenophophorea
- 4.4尿細管症
- 4.5グロボタラメア
- 5形態
- 5.1サイズ
- 5.2 - プロトプラズマ
- 5.3 - スケルトンまたはシェル
- 5.4 - 有孔虫のタイプ
- 5.5 - セドポッド
- 6ライフサイクル
- 7生殖
- 8栄養
- 9アプリケーション
- 9.1モーリシャスの場合
- 10参考文献
既知の種
現在では1万種以上が知られていますが、約4万種が絶滅しています。いくつかの種は海底を生息地として持っている、すなわち、それらは底生生物であり、しばしばエピファウナの一部を形成する砂の上に偽装されて生きている(epibetónicos)か、砂の下に住んでいることができる。このため、それらは生きている砂としても知られています。.
彼らは植物に住むこともでき、そこではそれらは着生植物として休み、そしてそれらの多くさえ固着生活を選択します、すなわち、彼らは彼らの存在を通して物質に固定されて生きます.
他の有孔虫は海のさまざまな深さ(0〜300 m)で浮遊していますが、それらは海洋性マイクロプランクトンの一部であるプランクトン生命です。これらの形態はより希少で多様性が低い.
最大かつ最も複雑な浮遊性有孔虫は熱帯や亜熱帯の環境でより一般的です。高緯度の環境では、これらの生物は通常乏しく、小さくそして非常に単純な方法で.
特徴
有孔虫で際立っている特徴は、絶滅した形態が海底に堆積する海洋微化石の形態で研究されたかもしれないという骨格または殻構造です。.
したがって、殻は有孔虫を区別するための基本的な要素であり、化石化する生物の唯一の構造です。これらの化石は海底堆積物中に非常に豊富にあり、堆積岩の形成にも関与しています。.
殻の主な化学化合物は方解石、アラゴナイトおよびシリカである。性器または無性生殖の産物であるかどうかにかかわらず、胚チャンバーの形状および寸法はその起源に依存する.
それらの個体発生の間に、有孔虫はチャンバーの成長と大きさを制御します。疑似足は鉱物殻に先行する有機膜の形成に関与するため、この制御は疑似足電流の長さおよび配置を通して行われる。.
カメラはバイオリアクターとして機能するため、このプロセスは細胞プロセスの維持に非常に重要です。.
有孔虫の大きさと形態に介在する要因
有孔虫が採用することができるサイズおよび最終形態は、以下を含むいくつかの要因に依存することに留意すべきである。
- 胚チャンバーの形状と寸法.
- 成体段階までの成長段階の数(すなわち有孔虫室の数).
- カメラの形状と個体発生中のその修正.
- カメラの配置.
大型の有孔虫は、最適なサイズを超えずに一定のチャンバーの容積を維持するための戦略的な成長パターンを持っています。これらの戦略は、クリークと呼ばれるいくつかのコンパートメントにカメラを分割することにあります.
これらのクラスターは、チャンバー内部の原形質と外部との間の輸送および調節機能を確実にするように配置されている。つまり、すべてのカメラとクリークは完全に相互接続されています.
室の配置は、直線または螺旋軸に従うことができる。これは、疑似足電流の位置およびチャンバ内の1つまたは複数の開口部の位置に左右される。.
分類法
ドメイン:真核生物
王国:Protista
範囲外:SARスーパーグループ
スーパーフィラム: Rhizaria
門: 有孔虫
クラスと注文
- アタラメア(網状粘膜)
- モノタラメア(同種異系、Astrorhizida、Komokiida)
- Xenophophorea(Psamminide、Stannomide)
- ツボタラメア(フスリニダ, インボルチニダ, ミリリオリド, シリコロクリニド, スピリリニド)
- グロボタラメア(Lituolida, ロフトシダ, シュルンヴェルリーニダ, Textulariida, Trochaminminida, ロタリド, ブリミニド, グロビゲリニド, Robertinida, カルテリニド, ラゲニダ).
分類
明確にすべきことはまだたくさんありますが、これまでのところ5つのクラスを区別できます。
アタラメア
ここであなたは殻を持っていないか裸である有孔虫を見つけることができます.
モノタラメア
それは単一の部屋で有機性か凝集した殻を提示するbetonic foraminiferaを含みます.
Xenophophorea
この場合、有孔虫は特大のBetonicタイプの大型、多核、および凝集殻を有する。それらは通常、腐敗性または腐肉食性です、すなわち、彼らは腐敗中の腐敗または有機物から彼らの食物を得ます.
ツボタラメア
これは、少なくとも幼若期に複数の管状室を有し、それが結合または石灰質の殻を有するらせん状に巻くことができる、ベトニック有孔虫を含む。.
グロボタラメア
この分類には、球状、凝集状、または石灰質の多殻性殻を持つ、ベトニックおよびプランクトンの両方の有孔虫が含まれます。シェルは、ユニセレート、biseriada、triseriada、またはtrocospiraladasにすることができます。.
しかし、この分類は絶えず進化しています.
形態学
-サイズ
有孔虫の大きさは通常0.1〜0.5 cmの範囲であり、いくつかの種は100μm〜20 cmの大きさである。.
-原形質
有孔虫は、有孔虫の細胞を構成する原形質塊によって形成される。.
原形質は通常無色ですが、少量の有機顔料、脂質材料、共生藻類、またはそれを着色する鉄化合物を含むことがあります。.
原形質は、小胞体と呼ばれる内部部分と外部部分の異所性からなる。.
小胞体ではそれは殻によって保護されており、その中では細胞小器官は消化液胞、核、ミトコンドリア、顆粒、ゴルジ体またはリボソームとして分布している。このため、それは粒状小胞と呼ばれることがあります。外形質は透明で、そこから格納式の偽足が出ています.
原形質は、ムコ多糖類の重ね合わせたシートによって構成される有機膜によって外部的に囲まれている。.
原形質体は、1つまたは複数の開口部(孔)を通って殻の外に広がり、外側に広がっています(カメラ外原形質)。これが偽足症の形成方法です。.
-スケルトンまたはシェル
有孔虫は、ミネラルスケルトン(殻)の構築により、細胞表面を恒久的に固定します。.
殻は隔壁によって分離された室によって構成されているが、同時にそれらは孔と呼ばれる相互接続孔を介して互いに連絡し、それ故に孔虫の名前である。骨格または殻の化学組成はそれらを非常に容易に化石化する構造にします.
部屋の内部はキチンと非常によく似た有機物で覆われています。さらに、シェルは主開口部を有することができる。毛穴を外側に持っていたり、欠けていたりすることもできます。.
鉱物殻は、単一区画(原始有孔虫またはモノタラモ)、または連続的に成長するチャンバー、または不連続成長の複雑な系における連続した段階で形成されるいくつかのチャンバー(有孔虫ポリラマ)によって形成することができる。.
この最後のプロセスは、以前に形成されたシェルへの追加、および戦略的な場所での新しい骨格材料の追加からなります。.
多くの有孔虫は、それらの化学組成、サイズまたは形状に従ってそれらの殻を形成するための材料を選択することができる。なぜなら、基材と接触している限界偽足電流はそれを認識することができるからである。.
-有孔虫の種類
シェルの構築形態によると、それらは3つの主要なタイプの有孔虫に分類することができます。
凝集した
この種の殻では、有孔虫は、それらが住んでいる環境で利用可能な大量の有機物質をそれらの偽嚢と共に集め、その後鉱物粒、スポンジの小穂、珪藻などのように凝集する。.
ほとんどの凝集性有孔虫は炭酸カルシウムで殻を固めますが、この化合物が環境中に存在しない場合、例えばカルシウムが存在しない海の深部に住むもののように、ケイ酸質、鉄質、有機セメントでそうすることができます。等
磁器
この場合、殻は有孔虫のゴルジ体で合成された方解石マグネシウム針を通して形成される。.
これらの針は、輸送されて外側に蓄積され、そして異物(セメント)のための接続要素として役立つか、または直接外部骨格を形成することができる。それらは高塩分環境(> 35%の塩分)で見つけられる.
彼らは通常無孔、すなわち、彼らは通常完全にシェルを通過しない疑似孔を持っています.
ヒアリン
これらは、原形質体の外部で行われるバイオミネラリゼーション(in situミネラリゼーション)と呼ばれるプロセスによって形成された有機テンプレートによる方解石結晶の成長によって形成されます。.
それらは壁の薄さのために透明であることを特徴とする。それらはまた気孔の位置、密度および直径が種に従って可変的であるところ穿孔されています.
-疑似足
この構造は動員、基質への付着、獲物の捕獲、そして骨格の創出のために使われます。偽脚の収縮および伸展のために、有孔虫は、多かれ少なかれ平行な列に配置された微小管の洗練されたネットワークを有する。.
疑似足の伸張は、体の長さの2倍または3倍に達することがあり、さらにその長さの最大20倍になることさえあります。これはそれぞれの特定の種によって異なります.
移動中の移動の種類は、シェルの形状と開口部の位置(疑似脚が由来する場所)に直接関係します。.
しかし、ほとんどの有孔虫は次のように移動します。疑似足は基質に付着してから細胞の残りを押します。このように移動すると、約1〜2.5 cm /時の速度で進むことができます。.
一方、有孔虫の偽足は、顆粒嚢の内部に顆粒を運ぶ双方向の細胞質内流動があるため、Granurreticulopodiaと呼ばれます。.
顆粒は、様々な材料の粒子、ミトコンドリア、消化液または排泄液胞、シンバイオティック渦鞭毛藻類などからなり得る。このため、このグループの同義語の1つはGranuloreticulosaです。.
偽脚のもう一つの重要な特徴は、それらが長く、細く、枝分かれして非常に豊富になりがちで、積み重ねることによって網状仮足のネットワークを形成する(吻合)ことである。.
ライフサイクル
有孔虫のライフサイクルは通常短く、通常数日または数週間ですが、大きな形態ではライフサイクルは2年に達することがあります.
期間は、有孔虫が受け入れるライフ戦略によって異なります。例えば、単純な形態を持つ小さい形状は短い日和見主義的戦略を開発する.
その一方で、シェルの大きなフォームと非常に複雑な形態を持つことは、保守的なライフ戦略を展開します。.
この最後の行動は、単細胞生物では非常に稀です。彼らは一様な人口密度と低成長を維持することができます.
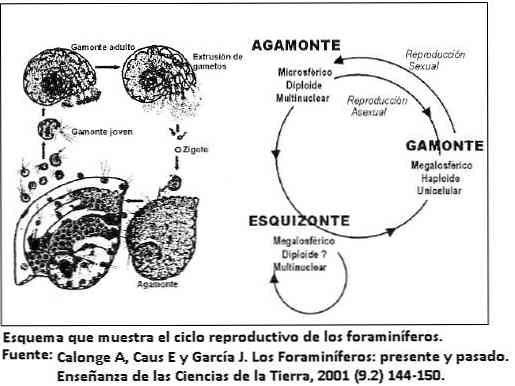
生殖
有孔虫のほとんどは、性的にのみ繁殖する浮遊性有孔虫を除いて、生殖の種類(性的または無性生殖)に応じた世代交代を伴う2つの形態を呈する。.
この形態の変化は二形性と呼ばれます。性的生殖(ガモゴニア)の結果として生じる形態はガモンと呼ばれ、無性生殖(分裂病)はシゾントの形態を与えます。どちらも形態学的に異なる.
有孔虫の中には、資源の利用を最適化するために、繁殖周期と季節周期を調整するものもあります。性的世代が奇形で起こる前に無性生殖が連続して起こるのを見ることは珍しくありません。.
これは、なぜシゾント形式がガモンテ形式よりも豊富であるかを説明しています。ガモンは、最初は単一の核を提示し、次に分割して多数の配偶子を生成します。.
シゾントは多核でありながら減数分裂後に新しい配偶子を形成する.
栄養
有孔虫は従属栄養性であることによって特徴付けられます、すなわち、それらは有機物を食べます.
この場合、有孔虫は主に珪藻または細菌を食べますが、他のより大きな種は線虫や甲殻類を食べます。囚人は彼らの偽脚を通して捕らえられている.
また、これらの生物は、緑、赤、金の藻類、珪藻類、渦鞭毛藻類など、多様な種類の共生藻類を使用することができ、同じ個体にそれらの非常に複雑な種類のものさえ存在する可能性があります.
一方、有孔虫のいくつかの種はクレプトプラストであり、これは摂取された藻からの葉緑体が有孔虫の一部となり光合成の機能を実行し続けることを意味する。.
これは生きるためにエネルギーを生産する代替方法を表しています.
アプリケーション
地質学的時間、進化、複雑さおよび大きさに沿った有孔虫の化石記録の豊富さはそれらを地球の現在および過去(地質学的時計)を研究するためのお気に入りの道具にしている.
したがって、その多様な種は、生層序、古生態、古海洋型の研究に非常に有用です。.
しかし、有孔虫集団の変化が環境の変化を示しているため、経済に影響を与える可能性がある生態学的災害を防ぐのにも役立ちます。.
例えば、殻のない有孔虫は、環境の変化に敏感であり、それらを取り巻く環境の変化に素早く反応します。したがって、それらは礁水の質と健康の研究のための理想的な指標種です。.
モーリシャスの場合
また、いくつかの出来事が私たちにこれを反映させました。これはモーリシャスで観察された現象の場合で、そこではビーチの白い砂の一部が消え、そして今彼らは観光客の流れを維持するためにマダガスカルからそれを輸入しなければならない.
そして、そこで何が起こりましたか?砂はどこから来るの?なぜそれが消えたのですか??
答えは以下の通りです。
砂は多くの生物の炭酸カルシウムの殻の蓄積、砂浜の端に引きずられる有孔虫に過ぎません。砂の消失は、炭酸塩生産者の漸進的かつ持続的な減少によるものです。.
これは、サトウキビなどの特定の製品を播種する際に肥料を過剰に使用しているために沿岸に達する窒素とリンで海が汚染された結果として起こりました。.
経済や社会に直接影響を与える、上記のような環境災害を防ぐために、社会科学で有孔虫を研究することが重要であるのはこのためです。.
参考文献
- ウィキペディアの貢献者. 有孔虫 [オンライン]ウィキペディア、フリー百科事典、2018年[相談日:2018年11月1日]。 es.wikipedia.orgで入手可能.
- Calonge A、Caus E、GarcíaJ. The Foraminifera:現在と過去. 地球科学の教え, 2001(9.2)144-150.
- チリ、ボカデルグアフォとゴルフォデペナス(43º-46ºs)の間のMicrobenthos(有孔虫:原生動物)のHromic T.生物多様性と生態. シエン。テクノール. 30(1):89−103,2007
- Humphreys AF、Halfar J、Ingle JC、他。ガラパゴスの低豊富な浅水底生有孔虫の分布と特性に対する海水温度、pH、栄養素の影響PLoS One。2018; 13(9):e0202746。 2018年9月12日公開doi:10.1371 / journal.pone.0202746
- De Vargas C、Norris R、Zaninetti L、Gibb SW、PawlowskiJ。浮遊性有孔虫における潜在的スペシエーションの分子的証拠およびそれらの海洋地域との関係。 Proc Natl Acad Sci US 1999; 96(6):2864−8.