シェア:



プラズマ学とは何ですか?
の 形質染色体学 それは配偶子または性細胞の細胞質の融合がそれらの核の融合なしに起こる、有性生殖の段階です。形質染色体性は真菌において一般的であり、有性生殖の第一段階である。それはまた融合および培養された植物および動物細胞においても起こり得る。.
配偶子は、その形態とそれらが果たす生殖機能によって、生物の他の細胞と区別される特別な細胞です。ある場合には、形質染色体性のプロセスは分化した配偶子の間ではなく、未分化の体細胞の間で起こる(体細胞性タイプの形質染色体性).

集中的な成長期間の後、菌類は繁殖期に入り、大量の胞子を形成および放出します。胞子は一般的に単細胞性であり、菌糸体の断片化によって、またはとりわけ胞子嚢、胞子体または配偶子などの特殊な構造内で産生される。.
胞子は、無性的にまたは間接的に有性生殖において産生され得る。真菌ならびに他の生物における有性生殖は、各個々の親の遺伝情報を含む2つの核の融合を含む。 2つの性細胞または配偶子が集まったときに核が物理的に見つかる.
索引
- 1真菌の有性生殖の段階
- 1.1プラズマ学
- 1.2カリオガミー
- 1.3減数分裂
- 2種類の形質染色体性
- 2.1配偶子の融合
- 2.2配偶者交尾
- 2.3ゲームマンギョの融合
- 2.4精子化
- 2.5ソマトガミア
- 3有性生殖の長所と短所
- 4参考文献
真菌の有性生殖の段階
有性生殖は、生物種の個体の遺伝的負荷を絶えず更新するメカニズムとして定義することができます。それは遺伝的多様性の重要な原因であり、それは新しい環境条件に適応するためのより大きな能力を可能にする。.
真菌の有性生殖のプロセスは、この王国のユニークで特別な特徴を持っています.
植物、動物、原生生物(分化した組織を含まない非常に単純な真核生物)のような他の真核生物(細胞膜や核小器官が膜で囲まれている)では、細胞分裂は核膜の溶解と再構築を伴う.
真菌では、核膜はプロセスを通して無傷のままです。例外であるいくつかの種では、核膜は壊れているが部分的にしかない.
真菌の有性生殖は、3段階で行われます:プラズマ染色体性、齲蝕性および減数分裂。各事象の持続期間または有性生殖の段階は異なり、これらの事象間の間隔も生物の種類に応じて変わります.
それほど進化していない原始真菌では、cariogamyは形質染色体性のほぼ直後に起こります。一方、より高く、より進化したキノコでは、両方の段階の間に間隔があります。.
形質染色体学
形質染色体または細胞融合は真菌における有性生殖の最初の段階であり、そこでは遺伝的に異なる一倍体細胞である2つの配偶子が併合され、2つの一倍体核を有する細胞が生じる。プラズマ学では、2つの一倍体の親配偶子の細胞質のみが結合する。.
半数体細胞は単一セットの染色体を含み、以下のように表される。 n. 二倍体細胞は2組の染色体または染色体の組を有する。それらは次のように象徴されています。 2n.
カリオガミー
キャリオガミーと呼ばれる次の段階では、親配偶子の2つの一倍体核の融合または結合が起こり、二倍体核を有する細胞が生じる。.
核の融合によって、接合子と呼ばれる新しい細胞が作り出されます。この接合体の核は、重複した数の染色体を含んでいます(すなわち、それは二倍体または2nです)。.
減数分裂
減数分裂は有性生殖の最後の段階であり、染色体の数は再び半分に減少します。減数分裂では、二倍体細胞(2n)は4個の半数体細胞(n)を産生する.
減数分裂では、新しい細胞の遺伝的構成(または遺伝的負荷)が全過程の前駆配偶子のそれと異なることを保証する染色体の組換え過程も起こる。.
プラズマ学の種類
真菌は、適合性細胞の2つの一倍体核を結合するために、すなわち形質染色体性が生じるために様々な方法を使用する。.
形質染色体性は、形態が異ならない細胞でより頻繁に起こり、この場合は同質性と呼ばれる。それらの細胞質を融合する細胞が異なるサイズのものであるとき、形質染色体はアニソガミーと呼ばれる。.
次のものがある5つの主なタイプの形質染色体学があります:配偶子の融合、配偶子の交尾、配偶子の融合、精子化および体性学。これらのタイプのプラズモグラフィーを以下に記載する。.
配偶子の融合
以前見たように、いくつかの真菌は、ガメタンギと呼ばれる性器から放出される特殊な性細胞(配偶子)を産生します。.
単細胞配偶子の融合は、それらの両方または少なくとも1つが移動可能である場合に起こる。胞子の可動性は、それらが泳ぐことを彼ら自身を推進することを可能にするべん毛を有することに依存し、その場合それらは動物胞子と呼ばれる。一般に、2つの融合配偶子は同じ大きさで、アイソガミック遊走子と呼ばれます。.
時折、1つの配偶子が他の配偶子(アニソガミック配偶子)より大きいことが起こるかもしれません。ジャンルでは モノブレファリス Chytridiomycota phyllaの、移動性の男性配偶子は男性の配偶子または子宮から解放されます.
続いて、雄の配偶子は雌の配偶子(oogoniumと呼ばれる)を貫通し、大きくて動かない雌の配偶子(oospheresと呼ばれる)を受精させる.
配偶者交尾
他の真菌では、2匹のガメタンジアが接触し、核がオスのガメタンギオからメスのガメタンギオへと通過する。この場合、gametangiosは配偶子の機能を果たします.
この種の形質染色体性はOomycotaグループの有機体で起こり、そこでは小さな雄の配偶子(anteridia)が成長し、枝分かれし、そしてより大きな雌の配偶子(oogonium)と融合する受精管を作る。.
受精管は、雄性配偶子の核が微細な貫通ピンを通過して雌性配偶子(卵球)と融合することを可能にする。.

ガムタンゴの融合
この種のプラズマゲームでは、配偶子は融合し、それらの核を結合する。例えば、Zigomycotaグループの真菌の胞子は形態学的に同一であり、一緒に成長し、そして接合体または卵を形成するように融合する分化した配偶子を形成する。この接合子は後で厚い壁の接合子胞子に変形する.
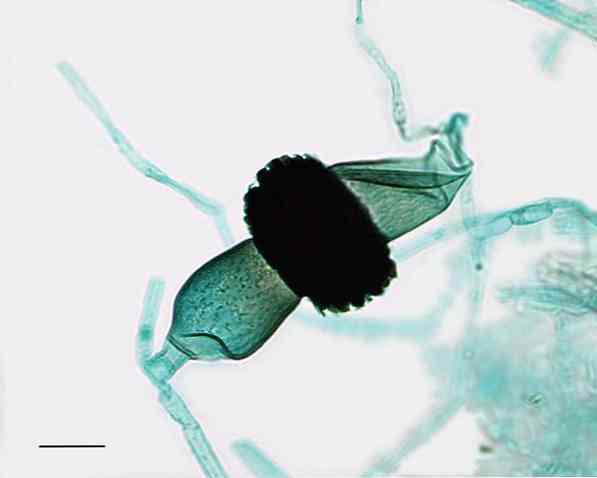
精子化
精子形成は、女性の配偶子と、単細胞(単核を有する)の、可動性(鞭毛なし)の融合からなる。.
ソマトガミア
いくつかのより進化した真菌は、配偶子を生成しません。これらの場合、真菌の体を形成する栄養体細胞性菌糸は、性機能を獲得し、接触し、互いに融合し、そしてそれらの核を交換する。.
この種の形質染色体性は、菌糸や酵母細胞のような栄養性のある非性的構造の融合で起こる.
有性生殖の長所と短所
有性生殖は、無性生殖と比較していくつかの不利な点があります。これらの不利益の中で、配偶子を作る際のより高いエネルギー消費、より遅い繁殖および結果としてのより少ない数の子孫を挙げることができる。.
一方、有性生殖は個人間で遺伝的変異を生じるという事実を利点として持っています。この種の繁殖では、子孫の遺伝的負荷は2人の親の遺伝子に由来し、これらのどれとも同一ではありません.
集団内の遺伝的多様性が大きいほど、その進化速度は速い。遺伝的変異性が高い集団は、それらが優れた適応能力を有する個体を産生することができるので、それらの環境の変化に対する異なる応答メカニズムを有する。.
参考文献
- Alexopoulus、C.J.、Mims、C.W。およびBlackwell、M.Editors。 (1996)。入門菌学第4版ニューヨーク:ジョン・ワイリーと息子.
- クラーク、J。およびハスキンス、E。 (2013)。粘液菌の核の生殖周期:レビュー。マイコスフィア4(2):233-248.doi:10.5943 / mycosphere / 4/2/6
- Dighton、J.(2016)。真菌生態系プロセス第2版ボカラトン:CRCプレス.
- Kavanah、K.編集。 (2017)菌類生物学と応用ニューヨーク:ジョン・ワイリー.
- アシュトンG.D.とDyer P. (2016)真菌の性的発達とその遺伝子発現系における使用In:Schmoll M.、DattenböckC.(eds)真菌における遺伝子発現システム:進歩と応用。真菌生物学スプリンガー.