シェア:



ウロコルダドス特性、分類、生息地、摂食
の ウロコルダドス またはチュニケート(subphylum Tunicata)は、もっぱら海に住んでいる非脊椎動物の脊索動物のグループです。それらのほとんどは、脊索が幼虫の尾部領域に限定されているので、それらはウロコルドードと呼ばれています。.
一方、名前tunicadosは、あなたの体がチュニカと呼ばれる多糖類の覆いによって保護されているという事実から来ています。ツニシンと呼ばれるこの多糖類は、セルロースに似た化学組成を持っています.
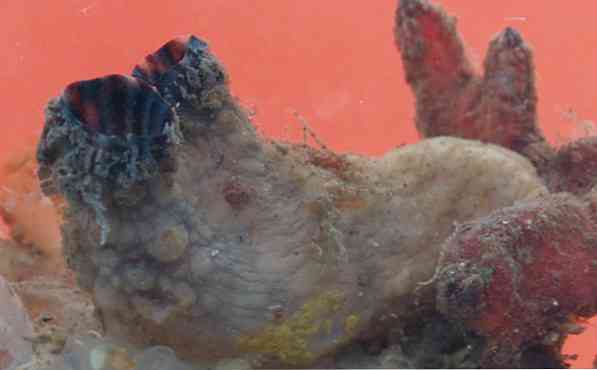
いくつかの種のチュニックは遠洋性ですが、ほとんどは底生生物です。彼らは一人暮らしや植民地を形成することができます。いくつかのホヤはまた構成されることができます、すなわち、何人かの個人が同じ吐き出すサイフォン(水がそれを通って有機体を出る構造)を共有するということです.
索引
- 1特徴
- 2分類と分類
- 2.1虫垂炎
- 2.2ホヤ科
- 2.3タリアセア
- 3生息地
- 4食べ物
- 5生殖
- 5.1無性
- 5.2性的
- 6重要性
- 6.1経済
- 6.2薬理学的
- 7参考文献
特徴
チュニックは和音であり、それゆえ彼らはそれらをそれ自体として定義する特徴を門の他のメンバーと共有します。これらは少なくとも胚期に存在し、そして:
-彼らはnotocordaと呼ばれる内部サポート構造を提示します。この構造は棒のような形をしており、結合組織の鞘に包まれた細胞で構成されています。それは、少なくともその幼虫期の間に、全ての脊索動物に存在する。.
-彼らは中空の背側神経索を持っています。神経コードを持つ無脊椎動物では、腹側に位置しています。脊索動物では、幼虫期の間に、それは消化管に対して背側に位置する。いくつかのグループでは、成人期に単純な神経節に縮小することができます.
-咽頭のえらスリットがあります。これらは咽頭と外部を連絡する開口部です。脊椎動物の羊膜(4つの胚体外膜を持つ胚発生を伴う四脚)では、これらの溝は開かないかもしれませんが、それらは単純な溝として形成され、残ります.
-内胚葉または甲状腺の存在も脊索動物の独特の特徴です.
urocordadosによって提示されたが他のコードではない他の特徴は以下の通りです。
-ツニシンで覆われた体.
-Notocordaは尾部のみで、通常は幼虫期のみです。しかしながら、虫垂外科医では、この構造は成人にも残る.
-消化管は「U」字型をしています.
-咽頭鰓裂の数が増加している.
-背側神経索は幼虫期にのみ存在する.
-彼らは2つのサイフォンを提示します。1つはサイフォンの吸入剤または発生と呼ばれる、もう1つはサイフォンの排出剤またはエクスカーションと呼ばれるその排出のためのものです。.
分類と分類
Tunicata分類群は1816年に、チュニックで覆われた体を提示した和音をグループ化するために、有名なフランスの自然主義者Jean-Baptiste Lamarckによって建てられました。 1881年に、イギリスの生物学者フランシス・メイトランド・バルフォアが同じ生物を分類するためにウロコルダタ分類群を建てました。.
おそらくBalfourの名声のために、グループの彼の分類は多くの科学者によって受け入れられました。長い間、両方の名前が異なる研究者によって使用されてきました。.
しかし、国際動物学命名法では、同様の場合、最も古い名前が優先されるべきであると定めています。この場合、Lamarckによって建てられた名前が優先されなければならず、したがって有効であると見なされる.
伝統的に、チュニックは4つのクラスに分けられました:虫垂炎(=幼虫)、ホヤ科、タリアシアおよびソルベラセア。この最後のクラスは1975年に建てられ、深海に生息するホヤのような有機体のグループを収容しました。.
sorberáceosは、以前はホヤの家族(Molgulidae)に分類されていました。その後、それらはヘキサクロビリダ科に移され、そこで彼らは階級レベルまで昇格するまで残っていました.
しかしながら、分子分析は、それらが示した形態学的な違いにもかかわらず、他のホヤ類とのそれらの近さを示した。このため、Sorberacea分類群は現在有効とは見なされていません。.
現在の分類によると、チュニケートの有効なクラスは次のとおりです。
虫垂炎
幼虫としても知られています。それらは浮遊性で孤独です、彼らはコロニーを形成しません。成体は、脊索や尾を含む幼虫の性格を保持しているので、彼らは新世代に苦しんでいると考えられています.
ホヤ科
それはグループ内で最も多様です。このクラスの代表者は、基質に固定して生きる底生生物です。それらは、孤独、植民地時代、あるいは複合的なものです。 2つのサイフォンは基板に対して反対方向に向けられている。神経コードは幼虫期に制限されています.
タリアセア
彼らはまた、サルップとして知られている遠洋習慣のチュニックです。サイフォンは反対方向に配置され、水泳で生物を助ける水流を生成するのに役立ちます。大人は尾を欠くが、えらスリットを保持.
生息地
チュニックはもっぱら海洋生物です。虫垂炎および距骨綱は遠洋性であり、一方、ホヤ科の子嚢(またはホヤ科)は底生生物です。その深度分布の観点から、それらは主に浅瀬に存在します、しかし、いくつかの種はもっぱら深淵です.
タリアセオスは赤道から極までのすべての海に住んでいますが、それらは暖かい水域でより頻繁にあります。同様に、彼らは浅瀬を好むが、いくつかの標本は1500メートルの深さで発見されている.
付録はプランクトンの一部です。彼らは孤独であり、自分自身によって分泌されるゼラチン状の構造に生息しています。彼らはすべての海の地表水にあります.
ホヤは固着性で、ほとんどあらゆる種類の基質に生きている。それらはすべての海と海に見られます。泥だらけの底に生息する種もありますが、それらは岩が多い基質でより頻繁にあります。彼らは潮間帯から深海の深さまで生息しています.
食べ物
チュニケートは主に濾過によって餌を与え、口腔または侵入サイフォンによって有機体の内部を貫通する水流を発生させ、それによってプランクトンおよび粒子状有機物の有機体を捕捉する。.
いくつかの種類の深海カモメは無脊椎動物を食べさせる捕食者です。彼らは口のサイフォンを使って、彼らに触れる獲物を捕まえる。他の種は泥だらけの基質の上に住んでいて、底に存在する有機物を食べます.
生殖
無性
それはtaliáceosとホヤで起こります。この種の繁殖は出芽によって起こります。 2種類の出芽が認められている:繁殖と生存.
伝播の発芽
通常、環境条件が適切であるときに発生します。ホヤの場合、それは基質の急速な定着を助けます。コロニーのサイズを増やすのにも役立ちます.
サバイバル発芽
環境条件が悪くなると、植民地は命名された芽を生み出します。これらは条件が悪い間は成長しません。状況が良くなると、芽は急成長します.
性的な
ほとんどのチュニックは同時雌雄同体です(つまり、個体は男性と女性の器官を同時に持っています)。ホヤでは、受精は外的または内的であり、オタマジャクシと呼ばれる幼虫に孵化する卵を作ります。しかし、いくつかの種では発生は直接的であり、それは幼虫期がないことを意味します.
タリアセオでは、ホヤのように、自由生活の幼虫がいない、性と無性世代の交代を示す種があり、有性生殖の間に内部受精を示す.
虫垂炎は性的繁殖のみを示していますが、これらの中では受精は外的です。それらは幼虫の発達を示し、生物は幼虫の特徴(ネオテニア)を維持しながら成熟する.

意義
経済的
ホヤの消費は非常に局所化されていますが、一部の国、主にアジア人では、これらの生物は非常に望ましいです。韓国では、種 Halocinthya roretzi それは栽培目的に使用され、2000年の売上高、18百万ドルを超える収益を生み出します.
近年、薬理学的に重要な生物活性物質の生産の可能性のために、作物および漁業の両方におけるこれらの生物の生産への関心が高まっている。.
それとは反対に、他の種のチュニケートは潜在的に有害である。基質をコロニー形成するその高い能力のために、いくつかの種のホヤは、二枚貝作物、主にカキおよびムール貝において害虫となる。.
薬理学的
ツニカートは、製薬産業に大きな可能性を秘めた多数の物質を生合成することができ、その中には、線状および環状ペプチド、アルカロイド、テルペノイド、ならびにイソプレノイドおよびヒドロキノンがある。このため、今世紀の初めまでに、全海洋天然物の5%以上がチュニックから来ました。.
ツニカートから得られる化合物の特性の中には、腫瘍細胞に対する中等度から高度の細胞毒性があり、それらはまた、抗可塑剤および抗トリパノソーマ活性を有することが示されている。.
海洋アルカロイドであるLepadinasは、パーキンソン病およびアルツハイマー病に関連するアセチルコリンのニューロン受容体に対して活性を示した。抗菌性、抗真菌性、抗ウイルス性、抗がん性、免疫抑制性および免疫刺激性を持つ物質も単離されています.
参考文献
- M.Tatián、C. Lagger、M. Demarchi&C. Mattoni(2011)。分子系統学は、肉食性およびフィルタ摂食性のチュニック(Tunicata、Ascidiacea)間の関係を支持する。スクリプトタ動物園.
- C.P.ヒックマン、L。 Roberts&A. Larson(1997)。動物学の総合原理マサチューセッツ州ボストン:WCB / McGraw-Hill.
- P.カストロ&M.フーバー(2003)。海洋生物学第4版、McGraw-Hill Co.
- R.C. Brusca、W. Moore&S.M.シャスター(2016)無脊椎動物第3版オックスフォード大学出版局.
- R。ロシャ、E。ゲラ - カストロ、C。リラ、S。ポール、I。ヘルナンデス、A。サレディ、J。ペレス、C。エレレア、A。カルボリーニ、V。カラバロ、D。サラザール、 M.Diaz&J.Cruz − Motta。 2010.ベネズエラのマルガリータ島のLa Restinga国立公園からのホヤ(Tunicata、Ascidiacea)の目録。ネオトロピカ生物相.
- J. Blunt、W. Copp、M。Munro、P。Norticote、およびM. Prinsep(2006)。海産物天然物ジャーナル.
- J. Petersen(2007)。ホヤ懸濁飼料実験海洋生物学および生態学.